|

|
|
|
Excursion du 22 Mai 1910 - Saint-Mandrier, Les Sablettes, Tamaris
Pour la premičre fois, l'excursion a été favorisée par un temps splendide. Dčs l'arrivée ą Saint-Mandrier, on gagne la cōte nord-est de la presqu'īle oł la récolte commence aussitōt sous la conduite de M. A. CHARLON.
Les botanistes recueillent dans les terrains vagues prčs de la mer : Glaucium flavum, Frankenia lœvis, Malva parviflora, Vaillantia muralis, Hyoscyamus albus et la forme H. major, Euphorbia Pithyusa.
Sur la falaise, prčs du Chāteau Dussaud : Helianthemum
Tuberaria, Linum gallicum, Tolpis barbata, Asterolinum stellalum, Coris
monspeliensis, Erythręa maritima, Globularia Alypum, Serapias cordigera, et les plantes parasites suivantes : Orobanche cruenta sur Coronilla juncea, Cytinus Hypocistis sur Cistus monspeliensis et C. salvifolius, ainsi qu'un pied de la variété rouge Kermesinus sur C. albidus, Limodorum abortivum sur les racines des pins.
Au bord de la mer, parmi les roches, sur le sable courent nombreux les Talitrus saltator.
Ces crustacés assez communs, sont parfois phosphorescents le soir ; ils
doivent cette particularité ą un microbe qui pullule dans leurs tissus.
On découvre en outre baignant dans la mer, un roseau garni d'une
colonie d'anatifes, Lepas anatifera.
En remontant la cōte, M. A. CHARLON fait remarquer dans une couche de haut permien un filon horizontal de barytine d'environ six centimčtres d'épaisseur qu'ila retrouvé sur toute la cōte de Saint-Mandrier ą Fabrégas.
Aprčs le déjeuner on se dirige sur les Sablettes ą travers bois. Sur les deux versants dominant le Creux Saint-Georges, se trouve la seule station franēaise du Julodis Onopordi F., beau bupreste qui habite aussi l'Espagne et l'Algérie ; dix sujets sont pris. Dans les galets de la plage, M. BAIZET capture Atelestus Peragalloi P., malachide brachélytre aptčre, spécial aussi ą la presqu'īle et, sur la plage des Sablettes, de nombreux Ateuchus semipunctatus.
On trouve sur le sable quelques coquilles gastéropodes : Cerithium vulgatum, Euthria collea, Natica cruentata, Astralium rugosum, Haliotis tuberculata et pélicipodes: Lima hians, Lima inflata, Cardium edule.
A 5 heures, M. C. VERLAQUE fait fort aimablement ą la Société les honneurs de sa serre de Tamaris, vrai bijou d'installation moderne contenant une trčs belle collection d'orchidées et de fougčres dont il serait difficile de citer toutes les richesses. Parmi celles en fleurs ą cette époque, il suffira de nommer cinq espčces de Cattleya, trois de Cypripedium, autant de Phalœnopsis, des Nepenthes, Selenipedium, Lœlia, Dendrobium, Œrides, Miltonia, un superbe hybride Lœlio-Cattleya, Cantramiana var. alba et un fort beau choix d'Oncidium. Autour contrastant par la légčreté de leur feuillage, d'élégantes fougčres : Adiantum, Gymnogramme, Nephrolepis, etc. Il convient de mentionner aussi les trčs curieux Platycerium, grande et alcicor qui vivent en épiphytes sur des plaques de ličge.
Coléoptčres nouveaux, rares ou intéressants pour le Département du Var, par M. R. MOLLANDIN DE BOISSY
Apalochrus flavolimbatus Muls. — Hyčres ! sur les phragmites dans le marais en été ; avait été pris autrefois en nombre aux Sablettes par l'abbé THOLIN.
Holoparamœcus Bertouti Aub. — Saint-Mandrier ! (BAIZET) sous les galets des plages ; rare.
Lixus flavescens Boh. — (verus).
Dans les collections, cette espčce est presque toujours confondue avec
d'autres ; il vit sur des Salsolacées halophiles (Sueda splendens) sur
lesquelles je l'ai pris aux Sablettes et aux Pesquiers ; ą l'état
frais, il est entičrement recouvertd'une exsudation jaune clair trčs dense.
OISEAUX
CRUSTACÉS.
MOLLUSQUES
(Extrait des Annales N° 1 - 1910).
M. GAIGNEBET raconte la visite de la Station de Biologie Marine de Tamaris le 3 octobre 1948. 35 participants. Le beau bātiment sinistré a été réparé grāce ą l'énergie de M. le Professeur Cordier, fermement appuyé par le Recteur de l'Université de Lyon M. Allix. Déją cet été, des étudiants et des professeurs franēais et étrangers ont pu y źtre accueillis. Une étude spécialement consacrée ą cette Station, ą sa fondation par notre regretté collčgue Raphaėl Dubois, ą son installation et ą son fonctionnement sera publiée dans nos Annales.
Une étroite liaison entre cet établissement scientifique et notre Société sera profitable ą chacun d'eux. M. le Professeur et Madame Cordier offrent une collation aux visiteurs qui se retirent enchantés.
(Extrait du Bulletin N° 29 - Décembre 1948).
Le 22 juillet 1956, une trentaine de membres de la Société ont visité la station biologique de Tamaris-sur-Mer, dirigée par M. le Professeur CORDIER. L'éminent Professeur de Physiologie leur donna des explications détaillées sur la station oł viennent travailler, chaque été, plusieurs savants et étudiants. Nous renouvelons nos remerciements trčs sincčres ą Mme et M. le Professeur CORDIER pour leur si cordial accueil.
(Extrait du Bulletin N° 98 - Octobre 1956).
Le Dr ANDRÉ, Président, a le regret d'annoncer le décčs (21 Octobre 1960) de M. Charles CORDIER (1900-1960), Professeur de Physiologie ą la Faculté des Sciences de Lyon, membre d'honneur de la Société et auteur de travaux remarquables. C'était un homme d'une amabilité extrźme, accueillant toujours avec courtoisie les membres de la Société, dans son laboratoire de Tamaris oł il venait tous les étés.
(Extrait du Bulletin N° 131 - Novembre-Décembre 1960).
Périodicité des composantes floristiques et faunistiques des peuplements marins du port de Saint-Elme (Var), par le Professeur Étienne GADEA.
I. - PRÉAMBULE.
J'ai étudié les peuplements marins du port de Saint-Elme, sous l'angle écologique et sociologique, au cours des années 1966 et 1967. Ce travail s'intčgre dans la progression de recherches dirigées par le Professeur Roger MOLINIER, de la Faculté des Sciences de Marseille, et relatives aux écobiocénoses marines superficielles de la Méditerranée. Il se situe plus précisément dans une succession d'analyses monographiques consacrées aux ports varois et animées par P. VIGNES dans le cadre de sa préparation de thčse de Doctorat d'État. Il constitue certainement, pour ma part, l'amorce de travaux ultérieurs qui s'orienteront cependant dans une voie sensiblement différente.
Dans leurs grandes lignes, les principes et méthodes de telles recherches ont été énoncés antérieurement (cf. bibliographie). Des relevés trimestriels approfondis, mais limités ą quelques stations, sont complétés par des observations plus fréquentes et plus étendues qui permettent d'embrasser la physionomie globale des peuplements. Des représentations figurées s'ajoutent aux tableaux de relevés sociologiques qui groupent - selon des notations conventionnelles - les résultats des divers recensements. Leur dépouillement procure de multiples enseignements, notamment sur la répartition des composantes - problčme ą reprendre ultérieurement - et leur périodicité, unique objet du présent article.
Pour bien connaītre un milieu, il faut le considérer sous tous ses aspects. L'écobiocénotique nécessite une triple enquźte visant : le MILIEU, la FLORE, la FAUNE. C'est sur ce découpage que s'appuiera directement mon exposé.
II. - LES CONDITIONS ÉCOLOGIQUES ET LEURS VARIATIONS SAISONNIČRES
Le port de Saint-Elme se situe sur la cōte varoise, ą l'extrémité sud-est de l'isthme sableux de la presqu'īle de Saint-Mandrier, dans le prolongement de la plage des Sablettes. La configuration du port, représenté ici tel qu'il fut étudié, (Planche I) est actuellement modifiée par suite de la modernisation des aménagements entreprise en juin 1967. Mais il s'agit toujours d'un abri de superficie modeste (0,5 ha environ) et de faible profondeur.
 |
Pźcheurs, plaisanciers et promeneurs, y entretiennent une activité permanente, modérée en hiver, beaucoup plus intense ą la belle saison. Cette influence humaine interfčre avec les conditions écologiques naturelles, constituant un complexe écologique au sein duquel nous distinguerons des facteurs physiques et des facteurs chimiques.
1.) - Les FACTEURS PHYSIQUES exercent une action énergétique qui se manifeste sous trois formes différentes : mécanique, photique, thermique.
a) L'agitation hydrodynamique.
Ą l'intérieur du bassin, le « mode » est calme, mais les vents dominants y suscitent des courants plus ou moins vifs et de direction différente. Ą l'extérieur des digues, l'effet mécanique des vagues et des courants est également lié ą la fréquence et ą la vitesse des mźmes vents. Les vents du sud et de l'est produisent une houle réfléchie qui entraīne une élévation du niveau. Le mistral pousse des lames courtes et rapprochées.
b) La lumičre.
L'éclairement, par son intensité, sa durée quotidienne et sa composition spectrale, conditionne les manifestations des organismes vivants, en premier lieu et directement celles de la flore. La durée quotidienne d'insolation croīt progressivement de janvier ą juin, puis diminue réguličrement de juillet ą décembre. Dans le milieu portuaire, les matičres en suspension, abondantes ą certaines époques, absorbent une fraction importante de l'énergie lumineuse. La présence d'écrans naturels (hautes frondaisons algales) ou artificielles (bateaux, quais...) , permanents ou temporaires, réduit la quantité totale de lumičre directe, réfléchie ou diffuse, favorisant les espčces sciaphiles au détriment des organismes photophiles.
c) La température.
L'eau de mer se réchauffe et se refroidit moins rapidement et moins fortement que l'air (Planche II). Cependant, dans le milieu portuaire oł l'eau est superficielle, mal renouvelée, l'eurythermie domine. Les températures élevées, surtout nettes ą faible profondeur du fait d'une forte absorption des radiations de grande longueur d'onde, ainsi que les températures anormalement basses de certaines journées d'hiver, ont des répercussions plus ou moins accusées qui vont jusqu'ą l'élimination brutale des composantes les plus sensibles.
2.) - Parmi les FACTEURS CHIMIQUES, il semble que les plus importants soient les substances polluantes qui favorisent la présence des espčces thionitrophiles. La pollution croīt ą partir du printemps, époque de la remise en état des bateaux de pźche et de plaisance, accompagnée de l'écoulement d'eaux sales, de détergents, de peinture, d'essence. Elle culmine en été par suite de l'activité plus grande des pźcheurs locaux qui, ą leur retour, rejettent par dessus bord les surplus inutilisables de leurs récoltes, animaux et végétaux dont beaucoup sont voués ą une décomposition rapide.
Tous ces facteurs. subjectivement décelés et appréciés, demeurent justiciables de mesures et de dosages précis qui se concevront surtout dans quelques années, ą l'aide d'un matériel spécial actuellement ą l'étude. Mais il est déją certain que leurs fluctuations interviennent puissamment sur la périodicité de la flore et de la faune.
 |
III. - PÉRIODICITÉ DE LA FLORE.
Dans une station donnée, les espčces observées ne se rencontrent pas tout au long de l'année avec une fréquence égale et le mźme développement. Les oscillations saisonničres concernent aussi bien le nombre et le volume des espčces que leur biologie.
1.) - PÉRIODICITÉ FLORISTIQUE GLOBALE.
P. VIGNES a montré que le catalogue floristique des peuplements du port des Salins d'Hyčres s'appauvrissait en été et se diversifiait en hiver. Ce fait se vérifie également pour le port de Saint-Elme.
Mais il est intéressant de noter l'évolution saisonničre inverse du nombre des espčces d'une part, de la masse qu'elles totalisent d'autre part (Planche III). La biomasse est maximale en été lorsque le nombre des espčces est le plus faible. Il faut voir lą probablement une donnée fondamentale de la dynamique des peuplements marins.
2.) - PÉRIODICITÉ DES GRANDS GROUPES TAXINOMIQUES.
Du point de vue pondéral, les Phéophycées affichent toujours une prédominance trčs nette par rapport aux Chlorophycées et aux Rhodophycées. Avec une participation de 76 %, leur avantage paraīt plus évident ici que dans les autres ports étudiés oł elles viennent néanmoins en tźte.
Du point de vue du nombre des espčces, les rapports concurrenciels sont plus nuancés (planche III).
Les Phéophycées sont réguličrement présentes, grāce ą la permanence de certaines espčces. Mais, au printemps, quelques algues brunes comme Cutleria multifida et Dilophus ligulatus font une apparition fugace.
En hiver, les Rhodophycées sont représentées par un nombre relativement élevé d'espčces sciaphiles, tandis qu'au printemps et en été se développent quelques algues rouges plus tolérantes ą l'égard de la lumičre.
Les Chlorophycées demeurent les plus discrčtes, ne revźtant une importance de premier plan qu'au voisinage immédiat du niveau, c'est-ą-dire au-dessus de mes stations de référence.
 |
3.) - PÉRIODICITÉ DES ESPČCES.
Ą chaque saison, j'ai noté la présence paroxysmale ou exclusive de certaines espčces :
- En hiver : Bangia fuscopurpurea, apparue dčs l'automne ; Scytosiphon lomentaria dont le développement se poursuit au printemps ; Udotea petiolata, encore relativement épargnée par les épiphytes ; diverses Rhodophycées comme Antithamnion plumula, Chondria boryana, Rhodymenia ardissonei et Peyssonnelia squamaria.
- Au printemps : Ceramium ciliatum, C. gracillimum, Laurencia sp., Gastroclonium clavatum, se manifestant depuis la fin de l'hiver. Cutleria multifida et Dilophus ligulatus ne sont visibles qu'un mois ou deux. Ą la fin du printemps, dans les eaux calmes du bassin, se développe une formation nébuleuse d'algues libres, physionomiquement homogčne. Elle est constituée par des thalles filamenteux d'Ectocarpales et de Cladophorales mźlées.
- En été : Cystoseira stricta ą l'extérieur du port et C. fimbriata ą l'intérieur ; Halopteris scoparia ; Acetabularia mediterranea.
- En automne Cystoseira crinita et C. fimbriata ą l'extérieur ; Hypnea musciformis ; Rivularia mesenterica au-dessus du niveau.
Mes observations sont conformes ą celles de divers auteurs et concordent, pour l'essentiel, avec celles de P. VIGNES au port des Salins d'Hyčres. Une fréquentation assidue du milieu étudié, ą raison de trois inspections mensuelles en moyenne, m'a en outre permis de dresser un diagramme de phénologie des composantes les plus facilement identifiables (planche IV). L'épaisseur du trait est proportionnelle ą l'extension de l'espčce.
 |
IV. - PÉRIODICITÉ DE LA FAUNE.
Comme la flore, la petite faune portuaire est sensible au cycle des saisons. Tenu compte de sa vagilité - l'éthologie prolonge alors l'écologie - elle répond également aux rythmes journaliers et ą divers stimuli, notamment par des déplacements verticaux qui lui font déserter temporairement les stations choisies pour des relevés méthodiques. Aussi l'interprétation des chiffres obtenus ne doit-elle źtre opérée qu'avec la plus extrźme prudence, certaines tendances s'avérant plus apparentes que réelles.
Une deuxičme particularité de la faune, liée ą la notion de chaīne alimentaire et de perte de rendement, est de totaliser une biomasse trčs inférieure ą celle de la flore. Aussi, parmi les diverses méthodes possibles d'analyse quantitative, ai-je donné la préférence ą celle qui s'appuie sur le dénombrement des individus.
1.) - PÉRIODICITÉ FAUNISTIQUE GLOBALE.
Le total des animaux récoltés pour les cinq stations de 625 cm2 de surface chacune, ą l'exception des pontes et des formes rnacroscopiquement non décelables, marque un maximum en été avec 1 286 individus. La régression amorcée en automne (853) se poursuit en hiver (784). Le minimum se situe ą la fin de l'hiver ou au début du printemps. Au cours du printemps, le nombre augmente ą nouveau (980). Un histogramme traduit bien cette pulsation annuelle (planche V).
 |
2.) - PÉRIODICITÉ DES GRANDS GROUPES TAXINOMIQUES.
Un tableau de valeurs numériques d'une part, un diagramme d'autre part, expriment les fluctuations saisonničres des principaux groupes. Pour les Gastropodes, la régression - peut-źtre anormale - enregistrée au printemps et en été semble en relation avec la fréquentation plus active du port par les enfants qui raclent journellement les quais avec leurs épuisettes.
3.) - PÉRIODICITÉ DES ESPČCES.
Comme je l'ai indiqué, les déterminations faunistiques de mes prélčvements par des spécialistes ne sont pas encore achevées. Deux exemples suggestifs illustreront cependant la complexité des liens qui unissent les organismes ą leur ambiance :
Le premier est fourni par un petit crabe, Acanthonyx lunulatus, recueilli dans les touffes de Cystoseira stricta, au lieu désigné « station IV ». Ses pattes, terminées par des pinces, lui permettent de s'agripper solidement et de résister aux assauts des vagues, dans ce secteur de mode battu. Il est curieux d'observer, mais en été seulement, des femelles avec leur ponte au sommet des buissons de cystoseires. Il serait intéressant d'expliquer, ą l'aide d'appareils ą préférendums, ce comportement qui peut źtre dū ą un besoin spécial d'oxygčne, de chaleur ou d'aliments.
Le deuxičme est donné par un Annélide Polychčte, Platynereis dumerili, présent dans toutes les stations. Trčs tolérante, cette espčce se révčle cosmopolite. On sait que les Polychčtes présentent souvent, au moment de leur reproduction, des phénomčnes d'épitoquie. Le nombre et la taille des individus récoltés prouvent, dans le cas présent, que la reproduction s'est accomplie au cours de l'été. Plusieurs auteurs ont montré que la maturité sexuelle est soumise aux phases lunaires et que l'essaimage des heteronereis dépend d'un phénomčne photopériodique : si le ciel est couvert ą l'époque de la pleine lune, la ponte est différée d'un mois.
Ces deux exemples montrent que l'ambiance agit sur la périodicité des espčces animales, en intervenant par exemple sur leur aptitude ą se nourrir ou ą se reproduire.
V. - CONCLUSION
Mes recherches, localisées dans le port de Saint-Elme et dont je n'ai pu évoquer ici que les éléments restreints, devraient apporter une certaine contribution ą diverses disciplines.
Du point de vue systématique, lorsque les déterminations animales seront achevées dans les laboratoires spécialisés, elles enrichiront le volet biologique de la fiche d'identité de certaines espčces, parfois encore insuffisamment connues.
Du point de vue écologique, leur apport est inégal selon l'échelle envisagée.
- Elles confirment
1. - le fait d'un déterminisme ą la fois climatique, édaphique et biotique, de la chronologie et de la distribution des espčces, au sein de la biosphčre tout entičre. Ce fait est déją solidement établi par ailleurs. Sous l'angle chronologique, par exemple, les modifications périodiques du milieu et les aptitudes biologiques des organismes interfčrent, justifiant les fluctuations spécifiques qu'il est donné partout de mettre en évidence.
2. - l'originalité des milieux aquatiques en général, du milieu marin plus particuličrement, avec pour premičre conséquence la présence exclusive ou préférentielle de nombreux groupes taxinomiques, Polychčtes par exemple.
3. - l'originalité de la Méditerranée, liée notamment ą des conditions thermiques et photiques différentes de celles qui rčgnent sur nos cōtes de l'Atlantique, de la Manche et de la Mer du Nord. Elle explique, non seulement la présence d'endémiques comme Rissoella verruculosa, mais encore la chronologie modifiée d'espčces ą distribution plus lāche comme Padina pavonia.
- Elles devraient se révéler davantage constructives dans l'optique de l'écobiocénotique portuaire, surtout plus tard lorsque la confrontation des relevés de nombreuses localités différentes, apportera les bases statistiques indispensables pour une tentative de généralisation valable.
- En tant que monographie particuličre, elles soulignent la personnalité d'un milieu qui ne trouvera certainement nulle part sa réplique parfaite, du fait que ne seront jamais réunies des conditions strictement identiques, sous l'angle par exemple du contexte géographique ou de l'influence humaine. C'est sans doute l'aspect le plus attachant de telles recherches.
FELDMANN (J.), 1937. - Recherches sur la végétation marine de la Méditerranée. La cōte des Albčres. Thčse. Rouen.
MOLINIER (Roger), 1960. - Etude des biocénoses marines du Cap Corse. Vegetatio, Haag, 9, p. 121-312.
MOLINIER (Roger) et VIGNES (Pierre), 1966. - Les problčmes de l'Écobiocénotique dans les domaines continental et océanique. Journées d'Écologie de Marseille. Centre Régional de Documentation Pédagogique d'Aix-Marseille, 22 pages.
SICSIC (M.) et VIGNES (P.), 1966. - L'intérźt biologique des milieux portuaires. Ann. Soc. Sc. Nat. et Arch. de Toulon et du Var, N° 18, 16 pages.
VIGNES (P.), 1965. - Principes et méthodes d'étude des peuplements littoraux superficiels. Information Scientifique, N° 1, 1965, 7 pages.
VIGNES (P.), 1965. - Contribution ą l'étude écologique et phytosociologique des peuplements marins du port des Salins d'Hyčres (Var). L'Information Scientifique, N° 3, 1965, 21 pages.
VIGNES (P.), 1966. - Quelques problčmes relatifs ą la distribution des espčces en milieu marin. L'Information Scientifique, N° 2, 1966, 22 pages.
(Extrait des Annales N° 19 - 1967).
Distribution comparée de 20 dominantes du peuplement algal dans le port des Lecques et le port de Saint-Elme (Var), par Antonia CARVOU, Etienne GADEA, Pierre VIGNES.
La présente étude s'inscrit dans un programme de recherches écobiocénotiques marines dirigées par le Professeur Roger MOLINIER de la Faculté des Sciences de Marseille.
Elle entre dans le contexte plus spécialisé d'une entreprise d'exploration méthodique des milieux portuaires varois, animée par l'un de nous (P. VIGNES) dans le cadre de sa Thčse de Doctorat d'Etat, en préparation.
Elle s'appuie sur une partie des travaux personnels des deux premiers signataires, recherches localisées dans le port des Lecques (A. CARVOU) et le port Saint-Elme (E. GADEA) et qui sont exposées in extenso dans leurs mémoires respectifs de Diplōmes d'Études Supérieures (1967).
I. - PRINCIPES DIRECTEURS
Les notions exposées ici proviennent de la rencontre d'une méthode de travail et d'un certain type de milieux naturels.
- La méthode est celle des cartes de distribution d'espčces ą trčs grande échelle (P. VIGNES, 1966).
Elle a fait déją l'objet de plusieurs démonstrations, ą des fins de recherche ou de pédagogie, dans un certain nombre de petites « localités » terrestres ou marines. Elle permet de hiérarchiser, par rapport ą tel ou tel facteur écologique, les besoins relatifs d'espčces territorialement voisines, ou encore ceux d'une mźme espčce par rapport ą plusieurs facteurs. Pour ce faire, les cartes de distribution locale sont comparées deux ą deux et logiquement classées.
Les présomptions réunies n'ont qu'une valeur qualitative. Mais elles préparent utilement des missions ultérieures de mesures et d'enregistrements de longue durée, climatiques et édaphiques. Elles permettent de choisir ą bon escient les points critiques d'une localité oł des coups de sonde écologiques trouveront le plus leur justification. Elles représentent une formule d'attente pour les biotopes marins benthiques, tant que les instruments appropriés - notamment l'ÉCOMČTRE polyvalent actuellement ą l'étude - ne sont pas disponibles.
 |
|
|
- Les milieux naturels sont ceux que recréent paradoxalement les petits ports, par le jeu d'une sorte de prescription qui efface trčs vite le caractčre d'abord artificiel de leurs aménagements. En rčgle générale, la flore algale s'y développe sans subir d'intervention intentionnellement sélective de la part de l'homme. Malgré l'emprise du béton et le déversement fréquent d'immondices, les peuplements peuvent y źtre considérés comme beaucoup plus spontanés que dans nos vertes campagnes oł l'on laboure, taille, arrache, répand herbicides et insecticides, contrariant constamment les tendances dynamiques locales.
En outre, les milieux portuaires sont clairement et fortement architecturés. Ils réunissent des conditions écologiques ą la fois variées d'une station ą l'autre et nettes dans une mźme station oł elles demeurent homogčnes sur d'assez grandes longueurs.
On conēoit l'étendue des promesses d'une méthode suggestive et d'application rapide dans des champs d'investigation aussi complaisants.
II. - CHOIX DES EXEMPLES
Trois options étaient ą considérer, relatives l'une aux ports ą comparer, la seconde aux espčces ą mettre en parallčle, la troisičme au mode précis de représentation ą adopter.
- Choix des ports.
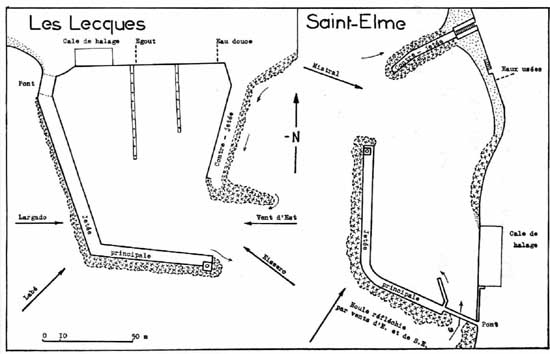
Onze ports varois ont fait l'objet jusqu'ą ce jour d'études écologiques et phytosociologiques axées sur la macroflore. Parmi eux. le port des Lecques et le port de Saint-Elme (figure 1) se prźtent ą une confrontation constructive.
Ils ont été étudiés la mźme année (1966) et les anomalies météorologiques de cette période n'ont pu que jouer dans un mźme sens sur leurs peuplements respectifs. Leur superficie est du mźme ordre de grandeur (0,8 hectare environ). Tous deux comportent une jetée principale coudée avec une section N-S et une section W-E. Ą l'origine de cette jetée, un pont fait communiquer les eaux libres avec celles du bassin en maintenant un courant qui s'oppose ą un envasement trop rapide. Peu profonds (&endash; 2 m dans le chenal d'entrée) les deux bassins n'hébergent que de petites embarcations de pźche et de plaisance, spécialement nombreuses en été. Les bateaux mouillent jusqu'au centre du plan d'eau oł ils sont accessibles, soit ą pied (Saint-Elme), soit au moyen de deux longs pontons (Les Lecques). Chacun des deux ports reēoit le débouché d'un égout dont le débit est maximal ą la saison touristique.
Certaines différences sont cependant notables. Le port de Saint-Elme est situé dans l'anse des Sablettes. Il est adossé ą la cōte par son cōté oriental. Sa contre-jetée est au nord et sa passe au nord-ouest. Le port des Lecques se trouve dans la baie de La Ciotat, ą plus de 20 km ą vol d'oiseau du précédent dont le séparent des accidents marqués de la ligne du littoral, le promontoire du Cap Sicié en particulier. Il s'accote au rivage par son bord septentrional. Sa contre-jetée est ą l'est et sa passe au sud-est.
- Choix des espčces.
Un effort de coordination a été tenté dans l'animation des diverses recherches portuaires varoises, Il était nécessairement limité, du fait que l'on peut préjuger jusqu'ą un certain point seulement de l'importance que revźtira en définitive telle ou telle composante du milieu étudié. En pratique, les peuplements ont donc été analysés d'une maničre relativement autonome aux Lecques et ą Saint-Elme, respectivement par A. CARVOU et E. GADEA qui n'étaient pas en rapport de travail direct et ne pouvaient se concerter. Or, sur 26 espčces cartographiées dans un cas et 36 dans l'autre, 20 espčces sont communes aux deux localités, celles-lą mźme dont les aires locales sont reproduites ci-contre (figure 2).
 |
|
|
Il ne s'agit d'ailleurs pas exactement de 20 espčces mais de 20 genres, espčces et formes. Sur 22 espčces, l'une d'elles a été dissociée en deux formes aisément reconnaissables in situ (Cystoseira fimbriata). Inversement, six autres espčces (Enteromorpha compressa et E. intestinalis, Laurencia pinnatifidea et L. undulata, Ulva lactuca et U. rigida) ont été regroupées au niveau générique : on ne pourrait les distinguer mieux ą coup sūr qu'ą la condition d'examiner un ą un tous leurs individus, opération trčs longue et de surcroīt nuisible ą l'intégrité des peuplements.
- Choix d'un mode précis de représentation.
Lorsque l'aire locale d'une espčce est trčs étalée en projection horizontale, elle peut et doit źtre concrétisée par une tache aux limites exactes. C'est ainsi que l'on procčde dans la plupart des milieux naturels.
Mais, dans les ports, l'aire d'une espčce est généralement linéaire parce qu'elle s'inscrit dans une zonation serrée et qu'en outre les aménagements en dur émergent étroitement de fonds meubles - ą l'intérieur comme ą l'extérieur - oł les algues ne peuvent s'établir durablement. Par souci de lisibilité, la distribution d'une composante est alors traduite par un liséré dont l'épaisseur est nécessairement excessive au regard de l'échelle cartographique réelle. On peut donc conférer ą cette épaisseur une signification symbolique, relative par exemple au degré d'abondance-dominance, ą la sociabilité, ą la vitalité ou encore ą une notion globale et subjective de « luxuriance ».
De mźme l'emplacement d'un point ou d'un tiret n'est pas l'affirmation de la localisation plane d'une touffe au mčtre prčs. La continuité ou l'intermittence du trait suggčre conventionnellement l'étirement ou la dislocation de l'espčce au sein de la ceinture de végétation qui l'héberge.
Il est encore prématuré de songer ą l'établissement de rčgles graphiques ą la fois précises et généralisables. Les membres de notre équipe ont procédé jusqu'ą présent, selon leurs tempéraments, d'une maničre intuitive ou selon des conventions personnelles mieux formulées, appliquées ą titre d'expérimentation méthodologique.
La représentation cartographique (figure 2) a été homogénéisée pour les besoins de cet exposé. Dans l'un des deux ports, les īlots de végétation étaient fréquemment figurés par des croix que nous avons remplacées ici par des points. Mais les remaniements des cartes se sont limités ą cette correction de pure forme.
 |
|
|
III. - INTERPRÉTATION ÉCOLOGIQUE
Un certain nombre de présomptions écologiques sont proposées dans le tableau ci-joint (figure 3), en regard des noms d'algues classés par ordre alphabétique. La vérité nous oblige ą dire qu'elles ne sont pas intégralement déduites des cartes monospécifiques des Lecques et de Saint-Elme. La fréquentation directe et répétée des milieux de référence a familiarisé A. CARVOU et E. GADEA avec des aspects de l'agencement des peuplements - zonation verticale en particulier - que les cartes ne restituent pas et pour lesquels conviendraient des figures complémentaires (profils, panoramas, perspectives). Une telle illustration n'a pu źtre retenue dans le cadre de la présente étude.
Par ailleurs les contacts assidus de P. VIGNES avec d'autres biotopes marins, portuaires ou sauvages ; ses notations personnelles dans des publications antérieures réalisées isolément ou avec la collaboration d'autres membres de l'équipe (E. BENSIMON, A. MEZE, M. PASTORE, M. SICSIC) ; l'expérience transmise dans leurs écrits par les grands écologistes marins (J. FELDMANN, Roger MOLINIER...) ; les communications verbales ; tous ces éléments de renfort sont venus étoffer notre échafaudage de quelques assurances supplémentaires. En toute éventualité, il n'y a pas de contradiction entre nos observations locales et les faits déją bien établis qui ne méritaient qu'un rappel. Mźme si des amendements ultérieurs sont inévitables dans le détail, l'ensemble de notre tentative paraīt cohérent.
Nous envisagerons successivement les effets de l'agitation, de l'éclairement, de la pollution. ainsi que d'autres circonstances écologiques non mentionnées dans le tableau.
- L'agitation.
Trois degrés ont été envisagés. Le signe +++ se rapporte ą la zone superficielle du secteur le plus battu, ą l'extérieur de la jetée principale. Le signe ++ concerne la zone superficielle des autres secteurs extérieurs, semi-battus. Le signe + convient pour l'intérieur du bassin ą tous les niveaux et les zones extérieures, plus profondes, de mode calme.
Il va de soi que, dans des biotopes différents, inobservables dans les localités prises pour exemple, le signe 0 (zéro) pourrait źtre affecté aux zones profondes tout ą fait épargnées par les mouvements oscillants de surface ; le signe ++++ traduirait une agitation trčs vive, telle que celle qui prévaut au niveau des bourrelets et trottoirs ą Lithophyllum tortuosum notamment.
L'agitation comporte des effets directs et des effets indirects.
°Effets directs : le facteur mécanique.
Rien ne prouve actuellement d'une maničre décisive que l'énergie cinétique ait une action physiologique favorable sur les espčces de mode battu. Il n'est pas impossible cependant qu'elle soit, chez certaines d'entre elles, un excitant de l'activité sécrétoire des cellules, bénéfique pour la croissance et le développement des thalles.
Il est certain, par contre, qu'elle est nuisible ą de nombreuses espčces qu'elle empźche de s'installer durablement. Seules résistent des algues adaptées, munies de dispositifs de fixation appropriés (disque adhésif de Cystoseira stricta, cellule basale renforcée par des rhizoļdes internes chez Bangia fuscopurpurea...) et que la forme et la consistance de leur thalle prémunissent contre l'arrachement : thalle coriace de Cystoseira stricta, filamenteux souple de Bangia fuscopurpurea, caoutchouteux de Nemalion helminthoides, en rosette courte et élastique des Laurencia, etc... Chez Cystoseira fimbriata, deux formes trčs distinctes traduisent une adaptation non héréditaire (ou accommodation), déją signalée par les algologues et maintes fois vérifiée dans les ports. Ą l'extérieur, en mode battu, c'est la forme en rosette courte dressée, trčs superficielle, présentant une convergence trompeuse avec Laurencia undulata. Ą l'intérieur, en mode calme, c'est la forme longuement rameuse, flottant ą l'aide d'aérocystes et implantée sensiblement plus bas.
°Effets indirects sur le facteur thermique.
Dans les zones battues, l'amplitude de variation des températures est faible. FELDMANN (1938) suppose que Cystoseira stricta doit ą son caractčre sténotherme sa relégation dans les biotopes superficiels battus. Il en est sans doute de mźme de Sphaerococcus coronopifolius que l'on observe non seulement dans des anfractuosités rocheuses relativement superficielles oł la turbulence des eaux est forte, mais aussi sur les fonds concrétionnés coralligčnes oł l'agitation de surface ne répercute plus d'écho et oł les températures demeurent ą peu prčs constantes. C'est un cas - parmi d'autres - de répartition bicéphale.
Le caractčre sténotherme d'espčces d'affinités froides comme Bangia fuscopurpurea et Nemalion helminthoides se traduit par leur distribution dans les secteurs battus mais aussi - parce qu'elles sont médiolittorales - leur disparition estivale complčte. Décalée dans le calendrier de phénologie, Acetabularia mediterranea d'affinités tropicales ne porte son disque en ombrelle qu'ą la belle saison.
Ą l'opposé, les espčces fortement eurythermes comme les ulves, les entéromorphes, prolifčrent toute l'année dans les bassins portuaires calmes, froids l'hiver, surchauffés l'été.
Certaines espčces de mode calme ou de mode semi-battu sont figurées sur les cartes dans des secteurs agités. Mais il convient de préciser qu'elles se localisent alors plus profondément. Le signe P est substitué au signe +++ sur le tableau récapitulatif dans tous les cas oł l'on observe un tel comportement.
- L'éclairement.
Les signes +++, ++ et + correspondent ą une échelle de valeurs non seulement subjective mais trop restreinte. Ils s'avčrent cependant commodes et se comprennent intuitivement, comme dans le cas de l'agitation. Ici encore on peut imaginer d'autres valeurs extrźmes, soit ++++ (eaux trčs limpides et forte réverbération) et 0 (nuit complčte en profondeur) qui ne se manifestent pas dans les deux localités prospectées.
Dans les petits ports, les composantes de la macroflore sont en majorité photophiles. En sous-strate se développe tout un cortčge de petites espčces plus ou moins sciaphiles qui ne sont gučre cartographiables présentement et dont nous ne ferons pas davantage état.
Les seules grandes espčces franchement sciaphiles représentées ici sont au nombre de trois : Udotea petiolata ą l'intérieur, Sphaerococcus coronopifolius ą l'extérieur, Dictyopteris membranacea de part et d'autre de la jetée. Cette distribution dissemblable tient ą l'interférence d'un mźme caractčre sciaphile avec des affinités inégales pour l'agitation. Le moindre éclairement résulte, soit de l'orientation générale du quai (Udotea, Dictyopteris pro parte), soit des accidents de la microtopographie, anfractuosités, surplombs (Sphaerococcus, Dictyopteris pro parte).
Des espčces photophiles se développent paradoxalement du cōté ombragé des jetées. Mais elles n'y occupent alors qu'une situation trčs superficielle, traduite sur le tableau par la lettre S.
- La pollution.
Il s'agit d'une notion écologique complexe. De l'activité humaine résulte l'accumulation de substances primitivement absentes ou ą l'état de traces dans les eaux marines : composés azotés rejetés par les égouts, composés soufrés libérés par fermentation d'ordures et de cadavres in situ, mazout et produits chimiques industriels plus ou moins accidentellement répandus.
Faible ą l'extérieur des jetées (+), la pollution devient forte dans les bassins oł les eaux sont peu renouvelées (++) et mźme trčs forte au plus profond des réduits portuaires ou face aux égouts (+++). Elle serait encore plus forte (++++) au débouché en mer de collecteurs urbains, ainsi que dans les grands ports ouverts aux bateaux de gros tonnage (Marseille, Toulon...). Les secteurs cōtiers oł elle s'annule pratiquement (0) deviennent l'exception.
Pour une espčce donnée, les matičres polluantes sont toxiques dčs que leur taux atteint un certain seuil, fréquemment dépassé localement de nos jours. Cystoseira stricta, par exemple, semble trčs sensible ą leurs méfaits comme l'indique sa régression au voisinage de Marseille.
Beaucoup d'espčces font preuve d'une tolérance plus grande et il est difficile, dans l'état présent de nos acquisitions, de préciser les conditions optimales de leur développement en fonction de la pollution. C'est pourquoi nous nous sommes généralement abstenus, sur le tableau, d'encadrer tel ou tel, signe en marquant par lą une préférence supposée.
Quelques algues enfin apparaissent comme positivement « thionitrophiles ». Ce terme proposé ą titre provisoire par l'un de nous (P. VIGNES 1965) correspond ą une certitude tout en constituant un constat d'ignorance. Ignorance car il est encore difficile de savoir lesquelles des matičres azotées et des matičres soufrées, dissociées au cours de la minéralisation bactérienne des protides, sont les plus déterminantes. Certitude car les espčces thionitrophiles atteignent un développement paroxysmal dans les secteurs les plus pollués des bassins portuaires, ą proximité des égouts notamment (+++).
Aux Lecques et ą Saint-Elme, c'est le cas pour les Ulva, les Enteromorpha et Scytosiphon lomentaria. Dans le premier port, ces algues occupent de concert tout le pourtour des deux longs pontons médians. Dans le second, elles coexistent sur l'épaulement rocheux qui borde sur une certaine longueur le fond du bassin. Dans notre optique anthropocentrique, il serait tentant d'en faire les parias des peuplements étudiés, au sein de sortes de ghettos phytosociologiques, végétant dans des stations déshéritées que les espčces concurrentes leur abandonnent. Mais le beau développement qu'elles y affichent exclut tout soupēon de misčre biologique.
- Autres circonstances écologiques.
Certaines données écologiques n'ont pas été méthodiquement recensées dans le tableau récapitulatif. Leurs variations ne sont pas étroitement calquées sur le tracé de la ligne de rivage et les besoins des espčces ą leur égard se déduisent mal - ou nullement - de la lecture des cartes monospécifiques. Quatre d'entre elles méritent cependant quelques commentaires.
°La nature physique du substrat.
De la consistance meuble ou compacte du substrat dépend l'efficacité du facteur mécanique sous l'angle de l'arrachage.
On redécouvre ą Saint-Elme et aux Lecques un tryptique habituel dans les petits ports et déją signalé ą propos des Salins-d'Hyčres (P. VIGNES 1966). Les algues s'installent aisément - bien que sélectivement en fonction du mode - sur les aménagements les plus superficiels en « dur », roche en place, rochers rapportés, béton. Plus bas, malgré la diminution de l'énergie cinétique des vagues, la végétation disparaīt sur les fonds sableux, vaseux ou jonchés de débris végétaux (litičre de posidonies). Plus bas encore, ą quelque distance des ports proprement dits, l'effet des vagues s'annule plus ou moins complčtement, ce qui permet l'implantation d'herbiers phanérogamiques.
L'existence d'un « no man's land » végétal explique l'étroitesse déją signalée ci-dessus, des liserés de répartition des espčces sur les cartes. Quelques exceptions qui se manifestent sur les cartes monospécifiques retiendront un instant notre attention.
Au sud du port de Saint-Elme, une plate-forme littorale gréseuse, accidentée dans le détail mais sans pente appréciable, brusquement terminée par une marche, héberge un certain nombre d'espčces de l'Ordre des Cystoseiretalia, Roger MOLINIER 1958. Cinq d'entre elles ont été cartographiées ici. (3 caractéristiques et 2 compagnes).
Ą Saint-Elme également, le bassin portuaire héberge de belles touffes de la forme longue de Cystoseira fimbriata, exprimée ici par conjonction du mode calme et de la dispersion de quelques cailloux ou tessons sur le fond.
Aux Lecques, les ulves colonisent une partie du bassin parce que des dalles rocheuses affleurent largement au milieu des sédiments.
Quant ą la tache massive de Falkenbergia rufolanosa dans le mźme port, elle correspond aux épaves d'innombrables individus de cette Rhodophycée épiphyte, mal prémunie contre l'arrachement, mais apte ą survivre longtemps, posée ou flottant prčs du fond dans les cuvettes calmes.
°La nature chimique du substrat.
On sait actuellement que la nature siliceuse ou calcaire du substrat est moins indifférente aux algues qu'on l'a longtemps cru. Certes, c'est dans leur ambiance liquide que les thalles puisent directement les substances minérales nécessaires ą leur métabolisme. Mais le phénomčne important de la fixation superficielle suppose des mécanismes chimiques de contact encore mal connus.
Le caractčre silicicole de l'endémique méditerranéenne Rissoella verruculosa (Observée ą Saint-Elme mais non aux Lecques et, par suite, non figurée dans cet article) est établi avec certitude. Les présomptions sont beaucoup moins avancées dans le cas général et nous ne nous étendrons pas davantage sur cette question, promise ą d'importants prolongements.
°L'eau.
Les espčces marines sont presque toutes aquatiques strictes. Le problčme de la résistance ą la déperdition d'eau ne se pose que pour les algues de l'étage médiolittoral.
Bangia fuscopurpurea et Nemalion helminthoides retiennent fortement l'eau par basse mer, du fait de leur richesse en mucilages. Si la dessiccation est cependant poussée, par suite d'une émersion prolongée sous un fort ensoleillement, ces algues se déshydratent fortement. Mais, douées de reviviscence, elles récupčrent en quelques instants le volume, la consistance, la coloration qui les caractérisaient en période d'activité.
Les Enteromorpha et Scytosiphon lomentaria se comportent différemment. Ą l'extérieur, en mode battu, elles sont médiolittorales dans des secteurs oł les vagues entretiennent leur humectation, mźme quand le niveau moyen de la mer est bas. Par mer ą la fois basse et calme, elles retiennent l'humidité par capillarité sous le tapis continu des thalles allongés, souples, ą forte sociabilité. Mais elles résistent mal ą une dessiccation poussée et prolongée.
Les mźmes algues deviennent franchement infralittorales, quoique superficielles, ą l'intérieur des bassins. Non seulement leur station diffčre et leur sociabilité diminue mais leur forme se modifie sensiblement. Les thalles deviennent plus largement tubuleux, s'enrichissent en sécrétions gazeuses ce qui favorise leur flottaison.
°Les chlorures.
Les algues marines sont en majorité sténohalines-halophiles. Quelques espčces cependant sont euryhalines-halophiles : les algues médiolittorales, comme Bangia fuscopurpurea, supportent aussi bien l'élévation du taux de salinité par évaporation ą marée basse que le dessalement sous les averses. Les espčces typiquement portuaires, ulves, entéromorphes, scytosiphons, admettent également des variations de chlorinité appréciables du fait des déversements d'eaux usées, de la dilution produite par les pluies d'orage ou, au contraire, des concentrations locales et temporaires, en été, dans le réduit le moins soumis au renouvellement des eaux.
CONCLUSION
Les études réalisées jusqu'ą ce jour par notre équipe permettent de tracer un « portrait-robot » d'un certain type de milieux portuaires. Il s'agit de ports petits ou moyens oł la pollution n'est pas excessive au point de devenir la donnée écologique primordiale - et limitante - et représente seulement un groupe de facteurs discriminants parmi d'autres. Par leur latitude, leur voisinage, ils s'inscrivent dans un mźme contexte climatique-régional et biogéographique. Les influences climatiques locales y font preuve d'une certaine constance : du fait de l'orientation générale des cōtes varoises, ils font face pour la plupart au sud (Les Lecques...), ą l'est (Saint-Elme) ou ą l'ouest (Saint-Pierre-d'Hyčres). Rares sont ceux qui sont tournés vers le nord (Saint-Mandrier).
Trois types de peuplements s'y observent :
- A l'extérieur, le spectre de zonation verticale est net et étalé. Les espčces médiolittorales y sont bien représentées. Celles des horizons supérieurs de l'étage infralittoral sont ą rattacher surtout ą l'Ordre des Cystoseiretalia, Roger MOLINIER 1958. Cystoseira stricta est le chef de file de ce groupe phytosociologique. Ses exigences (mode battu, sténothermie, fort éclairement, pollution minime) y sont bien satisfaites.
On observe cependant ą l'extérieur des jetées quelques espčces qui - pour ne pas źtre strictement inféodées aux milieux portuaires - ne semblent pas moins y trouver leurs conditions d'existence optimales et marquer pour la pollution plus qu'une simple tolérance. Tel serait le cas de Bangia fuscopurpurea et de Ceramium ciliatum qu'il est tentant de considérer comme des préférentielles semi-portuaires.
- A l'intérieur des jetées coexistent des espčces tolérant une pollution moyenne mais dont les besoins les plus accusés diffčrent. Les unes exigent avant tout un mode calme, d'autres un éclairement médiocre. Telles sont ici Cutleria multifida, Cystoseira fimbriata forme longue, Dictyota dichotoma, Dictyopteris membranacea, Udotea petiolata, etc... Ces peuplements hétérogčnes, variables d'un port ą un autre, ne peuvent dčs ą présent prendre place dans une mźme unité phytosociologique définie. Leur existence est néanmoins un fait général dans les petits ports. Il semble qu'ils n'aient pas la signification qui leur était d'abord accordée d'une « frange d'interférence » entre des blocs phytosociologiques en compétition ; qu'ils ne soient pas appelés ą glisser latéralement au cours du temps, dans le cadre d'une évolution dynamique qui amčnerait la suprématie progressive des éléments de l'un des camps principaux. Ce sont en fait des facičs de transition, constants dans la mesure oł la silhouette des ports demeure elle-mźme ą peu prčs constante.
- Au fond des réduits portuaires, la zonation verticale se ramčne ą sa plus simple expression. Une seule ceinture demeure souvent reconnaissable. Les peuplements sont affiliés ą l'Ordre des Ulvetalia Roger MOLINIER 1958, caractéristique des milieux portuaires. Les Ulva y apparaissent comme les chefs de file indiscutables. De mode calme, eurythermes et euryhalines, thionitrophiles, fortement photophiles, elles apportent ici la démonstration parfaite de leurs préférences et de leurs tolérances. Comme beaucoup d'espčces terrestres nitrophiles, elles tendent au cosmopolitisme. Les conditions chimiques localement réalisées semblent plus déterminantes que les conditions physiques et leur confčrent une relative indépendance ą l'égard des climats zonaux.
Il est permis en outre d'estimer qu'une Association végétale, caractérisée par des écotypes particuliers des ulves, des entéromorphes et des scytosiphons, accompagnée d'une faune spéciale, y réalise une biocénose originale. Cette biocénose fera l'objet d'une description prochaine quand ses éléments de diagnose seront plus serrés par le renfort de la microflore et de la microfaune.
Tels sont quelques-uns des enseignements tirés de la mise en pratique de la méthode de cartographie monospécifique dans les milieux de vie portuaires.
BENSIMON E., MEZE A., PASTORE M. et VIGNES P., 1966. - Rapports entre la forme et la distribution de quelques algues du littoral varois. - Annales de la S.S.N.A.T.V.
FELDMANN J., 1938. - Recherches sur la végétation marine de la Méditerranée ; la cōte des Albčres. - Revue Algologique, tome X, fasc. 1-4.
GADEA E., 1967. - Périodicité des composantes floristiques et faunistiques des peuplements marins du port de Saint-Elme (Var). - Annales de la S.S.N.A.T.V.
MOLINIER Roger, 1960. - Étude des biocénoses marines du Cap Corse. Vegetatio, volume IX, fasc. 3-5.
MOLINIER Roger et VIDAL M., 1963. - Études écologiques et biocénotiques dans la baie du Brusc (Var). Contribution ą l'étude du peuplement infralittoral de substrat solide dans le port du Brusc. - Bulletin de l'Institut Océanographique de Monaco, volume 61, n° 1273.
MOLINIER Roger et VIGNES P., 1966. - Les problčmes de l'écobiocénotique dans les domaines continental et océanique. - Annales du C.R.D.P. d'Aix-Marseille.
SICSIC M. et VIGNES P., 1966. - L'intérźt biologique des milieux portuaires. - Annales de la S.S.N.A.T.V.
VALLET G. et VIGNES P., 1967. - Les composantes floristiques, faunistiques, climatiques et édaphiques d'un milieu de vie naturel ; les effets écologiques de leurs rapports - Annales du C.R.D.P. d'Aix-Marseille.
VIGNES P., 1965. - Contribution ą l'étude écologique et phytosociologique des peuplements marins du port des Salins-d'Hyčres (Var). L'Information Scientifique, n° 3.
VIGNES P., 1966. - Quelques problčmes relatifs ą la distribution des espčces en milieu marin. - L'Information Scientifique, n° 2.
VIGNES P., 1966. - Étude écologique et phytosociologique du peuplement végétal d'un petit domaine. - Annales du C.R.D.P. d'Aix-Marseille.
VIGNES P., 1968. - Étude écologique de peuplements marins superficiels sur substrat rocheux. - Annales du C.R.D.P. d'Aix-Marseille.
N.B. 1 : Ą cette liste, il convient d'ajouter les mémoires des D.E.S. soutenus ą la Faculté des Sciences de Marseille-Saint-Charles par : VIDAL M. (1962), VIGNES P. (1964), SICSIC M. (1966), BENSIMON E. (1967), CARVOU A. (1967), GADEA E. (1967), MAGGI P. (1967).
N.B. 2 : Nous avons cité ici seulement les travaux concernant :
(Extrait des Annales N° 20 - 1968).
PROTECTION DE LA MER : Cartographie des fonds marins de la région de Toulon, par le groupe « ECOMAIR », par Jean-Marie ASTIER, Professeur au C.E.S. Bazeilles.
(...).
I. - Le groupe d'Écologie Marine ou « ECOMAIR-TOULON ».
1. - Son origine. Ainsi lors d'une séance de travail écologique tenue le 9 avril 19 7 3 ą la mairie de Toulon ą l'initiative de M. Henri Fabre, l'adjoint au maire, et du docteur Franēois Trucy, conseiller municipal délégué aux Espaces Verts, il fut créée une Commission extra municipale d'Ecologie marine. Le commandant Philippe Tailliez, membre du Comité Scientifique de Port-Cros, fut alors investi des plus larges responsabilités pour organiser, promouvoir, et rendre opérationnel un groupe d'écologie marine baptisé "Ecomair-Toulon" dont les activités s'inscrivent dans le cadre de la Commission extra municipale d'Ecologie marine.
Cette organisation ą caractčre purement civique, profitant des expériences acquises par 10 ans de campagnes océanographiques ą Port-Cros, poursuivrait, selon un programme ą long terme, des études écologiques localisées au milieu marin situé en regard de l'aire toulonnaise, de Sicié ą la presqu'īle de Giens.
(...).
III. - RECHERCHES ACTUELLEMENT EN COURS PAR LE GROUPE ECOMAIR
La cartographie sous-marine entreprise est une sorte de cadastre actuel des fonds marins ; c'est le point zéro pour savoir oł en est le patrimoine de la flore et de la faune. Dans une dizaine d'années, ces cartes permettront d'apprécier l'évolution des peuplements.
Avec tout son dynamisme, le lieutenant de vaisseau Jean-Marc Planēon a organisé les diverses Campagnes d'Océanographie de 1973, et a donné l'impulsion nécessaire pour intéresser et mźme passionner les plongeurs-démineurs ą des travaux sortant de la routine habituelle.
1. - En rade abri ą Toulon.
L'inventaire eut lieu en aoūt 1973, il fut effectué 44 radiales représentant 25 kilomčtres de longueur et couvrant 320 hectares qui ont permis la réalisation d'une carte au 1/5 000e. C'est le biologiste Pierre Maggi de l'Institut Scientifique et Technique des Pźches Maritimes de Nantes qui anima cette campagne océanographique et il fut amené ą constater que la baie de La Seyne et du Lazaret devait źtre encore recouvert, il y a une centaine d'années, d'herbiers de Posidonies, de Cymodocées et de Zostčres. La construction de la « grande jetée » de 1878 ą 1881 a transformé la rade abri en pičge ą vase et les Posidonies qui ne tolčrent pas des eaux turbides n'ont pu survivre. Les mollusques bivalves abondent dans la baie du Lazaret. Outre la mytiliculture cette zone se prźte parfaitement ą l'élevage de divers coquillages.
(...).
3. - Prospection des émissaires.
En collaboration avec la Cellule Antipollution du Var du Service de l'Equipement, le groupe ECOMAR a entrepris ą partir de 1974 une prospection du Benthos et sédiment en regard des 12 émissaires de rejets urbains du Cap Sicié ą la presqu'īle de Giens, afin de constater l'impact cumulatif des effluents sur les fonds marins depuis la mise en service de chaque émissaire, dresser l'état actuel des lieux avant prolongement éventuel de ces canalisations sous-marines.
Ą Sicié, Louis Del Buono, entrepreneur de ferronnerie, dirigea une prospection ą laquelle participčrent le club civil de plongée « Jules Corman » de La Seyne et la Société Nationale de Sauvetage en mer de Saint-Elme. Sept radiales permirent de circonscrire la zone des fonds marins dégradés par les effluents avant d'źtre inventoriée par les plongeurs-démineurs commandés par le lieutenant de vaisseau Masuy soucieux d'accomplir dans cette zone critique un travail de précision.

Les premičres séries des radiales de prospection (fig. 1) en regard du débouché du grand collecteur de Sicié montrent une disparition progressive de l'herbier de Posidonie au fur et ą mesure que l'on se rapproche de la cōte oł débouche l'émissaire, elles sont de plus en plus recouvertes d'Algues filamenteuses. On y trouve une abondance d'ophiures regroupées dans les creux de sédiments et des holothuries. Ą l'Est, les fonds, ą l'exception des holothuries, sont macroscopiquement azoļques, mźme ą &endash;35 m. Le sédiment est riche en un broyat renfermant morceaux de papier, peaux de fruits, graines, élastiques, cheveux... ayant échappé ą la dilacération et ą la putréfaction. La destruction des fonds est nettement plus importante ą l'Est de l'émissaire qu'ą l'Ouest. Une observation inquiétante a été faite : des poissons pélagiques, en particulier des poissons-aiguilles ou Orphies (Belone belone) ont été trouvés morts, au fond, ą prčs d'un kilomčtre de l'émissaire. Par contre aucun des nombreux Rougets de roche (Mullus surmuletus) qui fouillent le sédiment, riche en débris coquilliers, entre les touffes éparses de Posidonies ne fut trouvé mort. Il faut noter que la « nappe de pollution » (fig. 1 radiale R2) s'étend davantage en surface qu'en profondeur, le débouché de l'émissaire étant sur la cōte au niveau + 0,50 m. Les ceintures d'algues Rissoella verruculosa et Cystoseira stricta disparaissent sur les 2,4 km de frange cōtičre.
L'émissaire du Cap Cépet, dont la prospection a été commencée par les plongeurs-démineurs de la Marine nationale, débouche ą 57 m de profondeur. Il crache par intermittence des effluents qui s'élčvent jusqu'ą 6 m au-dessus du fond. On y trouve une biocœnose coralligčne fortement dégradée. Les gorgones mortes (Eunicella stricta) sont recouvertes d'algues filamenteuses, mais les bryozoaires rameux (Porella cervicornis, Tubucellaria cereoļdes) vivent encore. Le détritique cōtier ą Lithothamnion avec galets et graviers est trčs pauvre. Un poisson-lune (Mola mola) sans doute attiré par des eaux plus chaudes dormait au débouché de la canalisation.
(Extrait des Annales N° 27 - 1975).
Répartition des biocénoses benthiques en fonction des substrats sédimentaires de la rade de Toulon, par M. BOURCIER et Cl. NODOT, A. JEUDY DE GRISSAC, J. TINE.
M. MARVILLE a présenté cet important travail de chercheurs de la région, dont 2 sont membres de notre Société. Il s'agit de l'étude du macrobenthos de substrats meubles de la rade aprčs prélčvements ą la benne Petersen.
Les auteurs ont pu délimiter des zones de sédiments vaseux, de sédiments sableux, de vases terrigčnes cōtičres, de fonds détritiques cōtiers, chacune possédant sa biocénose particuličre.
Les espčces indicatrices de l'abondance de matičre organique existent dans toute la rade, surtout au N.E. oł le pélécypode Corbula (Aloides) gibba forme un facičs (3 600 ą 5 350 individus au m2). Les individus des espčces indicatrices de pollution ne sont pas trčs abondants, mais se rencontrent réguličrement dans les 2/3 de la zone Nord de la rade.
Il semble que les conditions naturelles de l'aire maritime toulonnaise aient été perturbées au cours des sičcles par les réalisations humaines : « Les fonds des baies Ouest (La Seyne) et Sud (Lazaret) étaient probablement occupés par des sables, vaseux en mode calme de type lagunaire protégés par un herbier de Posidonia oceanica... La partie médiane de la rade était occupée autrefois par des sables grossiers de type « ą Amphioxus ». La partie N.E. devait l'źtre par des sables fins bien calibrés et de hauts niveaux formant une vaste plage qui fut probablement le premier site d'implantation de la colonie toulonnaise ».
(Extrait du Bulletin N° 226 - 2e trimestre 1980).
Pour un plan d'occupation des fonds marins : inventaire des herbiers de posidonies du littoral du Var, par Jean-Marie ASTIER et Philippe TAILLIEZ
Estimation des superficies d'herbier de posidonies dans les six baies du département du Var (voir la carte) :
 |
BAIE 1 :
De la limite départementale Var-Bouches-du-Rhōne au Cap Sicié
908 ha
BAIE 2 :
Du Cap Sicié ą la pointe d'Escampobariou
1 989 ha
BAIE 3 :
De la pointe d'Escampobariou au Cap Bénat
7 954 ha
BAIE 4 :
Du Cap Bénat au Cap Lardier
1 964 ha
BAIE 5 :
Du Cap Lardier ą la pointe des Issambres
2 954 ha
BAIE 6 :
De la pointe des Issambres ą la limite départementale Var - Alpes-Maritimes
891 ha
TOTAL
16 660 ha
Destruction des herbiers par la pollution des égouts de l'agglomération toulonnaise
L'émissaire du Cap Sicié déverse les effluents non épurés des communes de Toulon, La Seyne, Ollioules, Six-Fours.
Le débit varie de 600 ą 800 l/s. Cet égout qui déverse les eaux usées juste au niveau de la mer a fortement perturbé les espčces fixées sur les fonds marins (peuplements benthiques) et détruit 75 ha d'herbier de posidonies.
Celui qui subsiste (3 ą 4 ha) est maladif et se situe ą 600 m de l'émissaire, vers 28 m de profondeur. La limite profonde de l'herbier forme une matte de 2 m de haut (Astier et Tailliez 1976 ; Astier et Tailliez 1978).
- L'émissaire de Sainte-Marguerite (Toulon Est, La Garde, La Valette) rejetait ses eaux usées par deux canalisations de 40 cm de diamčtre par 48 m de fond, ą 1 800 m du rivage.
Une station d'épuration a été construite. La partie des fonds les plus dégradés se situent ą l'Est, vers la baie de la Garonne.
- L'émissaire du Cap Cépet déverse les eaux non épurées de Saint-Mandrier et des bases de la Marine Nationale ą 57 m de profondeur. L'impact sur les fonds marins est moins important que celui de Sainte-Marguerite et touche le détritique cōtier, une biocoenose du coralligčne est fortement dégradée (Astier 1975).
- De nombreux ruisseaux et égouts pluviaux contribuent aussi ą la dégradation des herbiers en Grande Rade, les eaux étant fortement polluées lors des orages.
BIBLIOGRAPHIE
ASTIER J.-M. et TAILLIEZ P., 1976. - Impact des effluents du grand collecteur du Cap Sicié (égouts de Toulon, La Seyne et Six-Fours) sur la vie des fonds marins. Document publié par le groupe « ECOMAIR », Toulon, 27 p.
ASTIER J.-M. et TAILLIEZ P., 1978. - Les rejets en mer. Impact des effluents du grand collecteur du Cap Sicié sur la vie des fonds marins. Bull. Fondation Scientifique Ricard, 3 : 13-23.
(Extrait des Annales N° 36, fascicule 1 - 1984).
(Extrait des Annales N° 45, fascicule 2 - 1993).
(Extrait des Annales N° 50, fascicule 3 - 1998).
Retour ą la page d'accueil des archives de Jean-Claude AUTRAN
Retour ą la page d'accueil du site
|
